What is Translocation in plants with Mechanism?
The Hypothesis of Phloem Translocation
From the 1930s to the mid-1970s, the mechanism of phloem translocation was a subject of research. Now, one theory is generally accepted as the correct explanation for translocation. This theory called the pressure-flow hypothesis is favored by most plant physiologists and was proposed by E. Munch in Germany in 1926.
Laboratory Model for Munch’s Pressure Flow Hypothesis
The hypothesis is based upon a real, model that can be built in the laboratory. The model consists of two osmometers connected to each other with a tube. The osmometers can be immersed in the same solution or in different solutions, which may or may not be connected.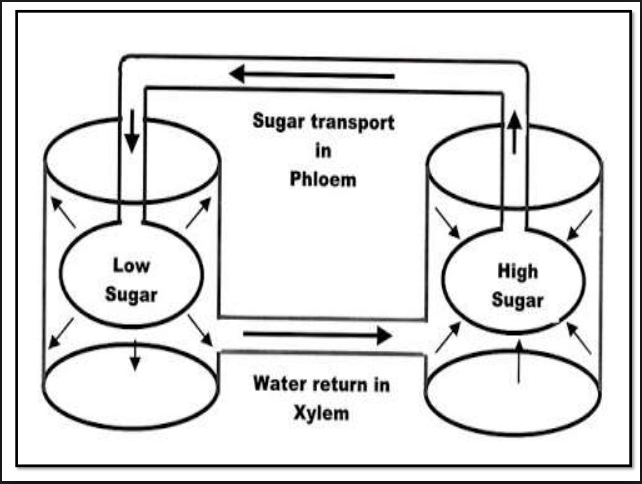
The first osmometer contains a solution more concentrated than the solution surrounding it, and then, the second osmometer contains a solution less concentrated than the solution surrounding it: The water moves into the first osmometer by osmosis and pressure builds in it. As the osmometers are connected, the pressure is transferred from the first to the second osmometer.
The pressure in the second osmometer increases so its water potential becomes more positive than in its surrounding solution. The water molecules diffuse out of the osmometer through the membrane into the surrounding solution whereas solute molecules are retained by the osmometer.
The net result is that water moves osmotically into the first osmometer from its surrounding solution, bulk flow of solution through the tube into the second osmometer, and osmotic movement of water out of the second osmometer into its surrounding solution.
lf the walls of the second osmometer stretch, the pressure is relieved even if no water moves out, and if the second osmometer is surrounded by a solution more concentrated than that inside it, water will diffuse into the surrounding solution even without buildup of pressure.
Equivalent System for Munch’s Pressure Flow Hypothesis
Munch suggested that the living plant contains a comparable system but with an advantage. Sieve elements near-source cells (usually photosynthesizing mesophyll cells) are analogous to the first osmometer, but the concentration of sugars is kept high in these sieve cells by sugars that are produced by photosynthesis in the nearby mesophyll cells.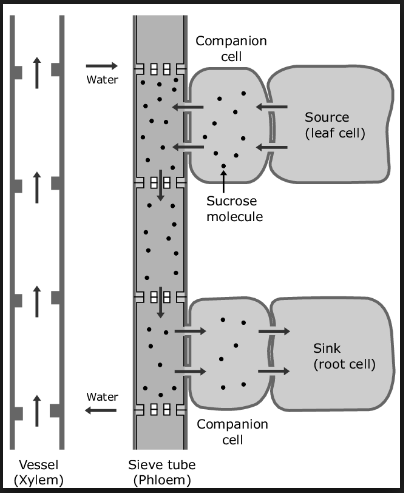
The concentration of sugars in the other end of the phloem system, near the sink cells is kept low as the sugars are utilized in metabolism, incorporated into protoplasm (growth), or stored ‘as starch or fats.
The metabolism, growth, and storage take place in cells near the sieve tubes in the sink tissues. The connecting channel between the source and sink is the phloem system with its sieve tubes; the surrounding dilute solutions are those in cell walls and xylem (apoplast).
Flow-through Sieve Tubes
The flow through the sieve tubes is passively occurring in response to the pressure gradient caused by osmotic diffusion of water into the sieve tubes at the source end of the system and out of the sieve tubes at the sink end of the system. There is no evidence of active pumping of the solution by sieve elements along the route.
However, active theories suggesting the involvement of energy are proposed to explain translocation in the phloem. According to one of the active theories, the solutes are carried from one end of a sieve element to the other by cytoplasmic streaming or cyclosis. But cytoplasmic streaming has never been observed in mature, functioning sieve elements.
When sugar is loaded by active transport into the sieve tube at the source, the total concentration of dissolved solutes will become greater than in adjoining cells. Therefore, water will move by osmosis from adjoining cells into the sieve tube.
Unloading of sugars by active transport at the sink results in the movement of water by osmosis from the sieve tube to adjoining cells. Thus, osmosis into the source and out of the sink establishes the flow of sugar solution in the sieve tube.
Phloem Loading (Phloem Translocation)
The active transport of sucrose into the sieve elements is called phloem loading. The process of transport inside the sieve elements from source to sink is called export.
Several steps are involved in the movement of the product of photosynthesis from the mesophyll cells to the sieve elements of mature leaves. These include:
- The sucrose synthesized in the leaf mesophyll cells passes to the minor leaf veins which may be adjacent to the mesophyll cell or 2 to 3 cells away from it.
- The movement within the mesophyll cells is from cell to cell through plasmodesmata (symplast).
- The sugar is actively transported out of mesophyll cells into the cell walls of the minor veins (apoplast).
- From the apoplast, the sugar is actively absorbed probably into the large companion cells of the minor veins from which it passes into the sieve elements through symplast.
Active loading of sucrose into the companion cells produces very negative osmotic potentials in those cells. This leads to an osmotic entrance of water, which then passes in bulk flow through plasmodesmata present between companion cells and sieve elements, carrying the sucrose along with it.
Thus the high concentration of sugar loaded in companion cells and then sieve elements and the resultant uptake of water by osmosis produces high pressures and mass flow in sieve tubes. In other words, phloem loading is responsible for the buildup of high pressures in sieve tubes.
Sucrose: Proton Cotransport Mechanism
Recent studies suggest that sucrose loading into the phloem occurs by a cotransport system. The protons are pumped out through the plasma membrane using the energy from the ATP and an ATPase carrier enzyme, so the pH outside the cell in the apoplast becomes much lower (more acidic) than inside the cell.
Protons then diffuse back into the cell, and their movement across the membrane is coupled to a carrier protein that transports sucrose into the cell along with hydrogen ion. Several pieces of evidence support the existence of such a cotransport mechanism.
For example, the contents of sieve tubes have relatively low ‘proton concentration and high potassium concentration. The pH at the loading site is quite acidic compared with the phloem sap (5.5. to 6.0). If the pH of the apoplast of a leaf is increased sucrose uptake is inhibited.
Phloem Unloading (Translocation of Sugar)
The process by which translocated sugars exit from the sieve elements of sink tissue is known as phloem unloading. Translocation into sink organs such as developing roots, tubers, and reproductive structures is called import. The initial step of the process is unloading from the sieve elements, and then the sugars are transported in the sink for storage or metabolism.
The Problems with Pressure Flow
The pressure-flow hypothesis visualizes a steady, unidirectional flow of sugar solution through living sieve elements. The flow is generated by osmosis. It involves continuous entry of sugar molecules at the source and exit at the sink by active transport, together with the entry of water by osmosis into the source and exit out of the sink. It helps in maintaining the flow of sugar solution along the length of the sieve tube.
The objection leveled against the pressure-flow hypothesis is that it does not explain the phenomenon of bidirectional movement, i.e., movement of different substances in opposite directions at the same time. The phenomenon of bidirectional movement can be demonstrated by applying two different substances at the same time to two different points of the phloem of a stem and following their longitudinal movement along the stem’ The bidirectional movement occurs in a single sieve tube or not.
If the mechanism of translocation operates according to the pressure-flow hypothesis, bidirectional movement in a single sieve tube is not possible. Experiments to demonstrate bidirectional movement in a single sieve tube is technically very difficult to perform. Some experiments indicate that bidirectional movement may occur in a single sieve tube, whereas others do not.
External Links……!
(https://www.bbc.com/bitesize/guides/zps82hv/revision/1)
(https://www.slideshare.net/eportch/translocation-in-plants)
(https://study.com/academy/lesson/plant-translocation-definition-mechanism.html)


Wow, awesome blog structure! How lengthy have you ever been blogging for? you make running a blog glance easy. The whole glance of your site is excellent, let alone the content material